Biology Forum › Zoology Discussion › Tyrannosaurus sex et alia topics
- AuthorPosts
- August 13, 2006 at 4:20 am #5474
 KiezelParticipant
KiezelParticipantI have an enigma for anyone able to assist thee…
Tyrannosaurus rex appears to have undergone a proximal-distal suppression of growth in the forelimbs where the arm apparently weakens the farther you travel down it, from the humerus to the phalanges. Make note that this supposedly is documented in lizards that are on the verge of becoming limbless. Now in the last decade or so, it has been made a big deal out of the fact that this particular tyrannosaurid could supposedly curl 400 + lbs in one arm. However, Bakker noted last year in our tyrannosaur symposium that the fingertips of this species could only produce a mere 0.2 horsepower in a spasm. Now of course if we were to leave it there, it would seem that these arms were vestigial. Another contemporary, the Edmontosaurus can apparently produce in a spurt a force of 35 horsepower around the rump region. The result of such an "athletic contest" is clear: those arms of our rexy are coming clean off in the next five seconds once it grabs its struggling prey.
BUT… AFAIK, there is a osteopathology found in almost, if not all, tyrannosaur humeri that have been recovered. The injuries are for the most part stress fractures, with one major injury accounted for in which the tendon attachted to the bone was ripped out of place, leaving a small depression and spur of bone on the humerus. What I find odd is why the forelimbs were apparently atrophying but still served some active purpose.
The most credible hypothesis I think that I have heard comes from Larson, Bakker et al, in which tyrannosaurs apparently had really, rough sex lives. This seems likely to me, as the same specimen with the badly torn humerus also seems to have sustained a nasty injury and regrowth of bone in the caudal vertebrae, possibly the poor bastard (pardonner mon français) stepping on his not-so-happy mate. I’m not necessarily suggesting the two palaeopathologies are connected to the exact same incident, but it appears to me as if they give support to the difficulty in mounting & copulation when your [female] partner is 10% larger than (or so the idea goes) you! Bakker has suggested male mounting and suppression as seen (and stereotypically characterized as sexual humping) in domesticated felines and canines. I seem to recall that such a display of dominance is also observed in komodos, but with mating intentions and occurring between male and female.
I am still befuddled by the use of the arms. Are there any modern examples of this sort of morphological transformation going on? Atrophying appendages that still serve a particular and specific use? My name is Kiezel by the way; jovial am I to be a member of this absorbing community.
Any thoughts? Feelings? Random outburst of emotion?
I’m curious as to the other opinions of such an intelligent audience.I beg pardon for my ramble
- August 14, 2006 at 12:33 pm #53344madnewtParticipant
I can think of no particular species in which appendages are being lost due to evolution and where that appendage is still being used.
I know very little about the T. rex so please forgive my ignorance, but are we sure that the front limbs are atrophying gradually over time or were they merely this weak at one particular point in time. If they were so important in copulation, it figures that those with the strongest forelimbs would on average give rise to the most offspring over the course of its life and thus evolution should encourage stronger limbs.
- August 14, 2006 at 1:38 pm #53347DarbyParticipant
Don’t some pythons have rear leg remnants that are used as graspers during copulation?
- August 14, 2006 at 6:55 pm #53375MrMisteryParticipant
Pythons do have leg remnants. But i have personally seen a dead python and i can tell you that the only way to see those remnants in a live python is to make an X-ray. I doubt they can be used for anything.
@madnewt
Good point.
And we know the front limbs of the T-rex were atrophied. this is one of the reasons why there have been so many suggestions that the T-rex was not even a predator, that he was a scavenger. - August 15, 2006 at 12:16 am #53391KiezelParticipantquote MrMistery:this is one of the reasons why there have been so many suggestions that the T-rex was not even a predator, that he was a scavenger.
True, but as far as I am aware, John Horner is the only paleontologist that really thinks small arms equate to a bad predator. The idea doesn’t hold strong for several reasons, the most obvious being that we have direct fossil evidence of tyrannosaur predation on the prey contemporaries. Concerning the arms, most extant and fairly active predators don’t use their arms to catch prey. Felines and primates predominantly use their forelimbs to snare prey. And yet canines, sharks, birds of prey, crocodiles and monitors et al all use either their mouths or –in the case of birds of prey– their feet.
Arms aren’t necessarily needed when you’ve got a four foot maw. I suppose, with the prevalent prey of tyrannosaurs exceeding 3 tons, that the preferred method of hunting would be ambush, with the predator throwing much of its bodyweight towards the broadside of the prey. Such an idea was also brought up last year too.
But going back to the topic of scavenging: it honestly seems as if Horner is using the publicity and the Discovery Channel to bring in funding. He seriously can’t support his idea of an obligate scavenging tyrannosaur anymore, all his ideas that he originally put forth to build his foundation have crumbled under the scrutiny of others. And the body plan for a obligate scavenger just doesn’t fit with tyrannosaurs. Of all vertebrates, only the 23 species of vultures are thought of as true, obligate scavengers, and it has been documented that even some of their own will kill too, during desperate times.
I’ll just bring up one last point. The hypothesis that tyrannosaurs were obligate scavengers because they are supposedly slow is completely arrogant of the osteological evidence. While tyrannosaurs may seem slow to us in a world filled with jet planes and Ferraris, it was certainly one of the fastest terrestrial animals for its size. Tyrannosaurids are characterized by long tibiae and metatarsi that bare a striking similiarity to the ostrich-like ornithomimosaurids. Dromaeosaurs, considered by Horner to be the better "runners" , have about the least gracile limb proportions of any theropod with the exception of therizinosauroids. Crichton and Horner aside, dromaeosaurids were probably not very fast runners relative to tyrannosaurids, ornithomimids, etc.

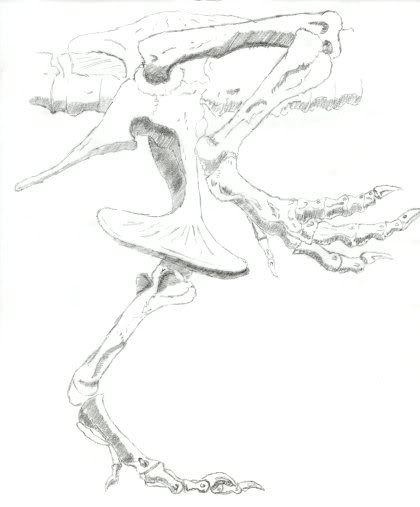
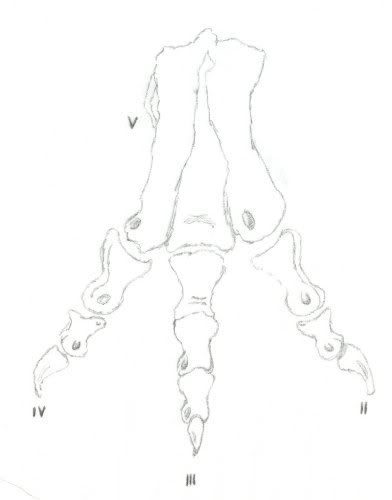
And like ornithomimosaurids, tyrannosaurids also have what is called an arctometatarsus. An arctometatarsus is where metatarsal III forms a wedge distally, but pinches out proximally. This feature is unique and found in some ornithomimosaurs, deinonychosaurs, alvareszaurids, and oviraptorosaurs. An arctometatarsus has also been shown to be a good and natural shock absorber for fast running.
Both sketches are by me for your viewing pleasure…
- August 16, 2006 at 1:11 am #53460DarbyParticipant
It was my impression that although all pythons have the rear-leg bits, only a couple of species have external, functional projections used to grip during copulation.
I’m not a fan of the obligate scavenger tyrannosaur, but how do you respond to the assertion that if they were fast runners, there would be more stress damage to the ankles-? (Which, oddly enough, I think comes from the same paleopathologist that was mentioned in the first post here).
- August 16, 2006 at 8:22 pm #53511MrMisteryParticipant
It’s not odd, that guy is the father of the theory. I mean hypothesis
- AuthorPosts
You must be logged in to reply to this topic.
No related posts.